
<English>
Penelitian oleh Dhani Irwanto, 27 Oktober 2015
Sundalandia adalah wilayah bio-geografi Asia Tenggara yang meliputi Paparan Sunda, bagian landas kontinen Asia yang terekspos selama Zaman Es terakhir. Periode Glasial Terakhir, dikenal sebagai Zaman Es Terakhir, adalah periode glasial terakhir dalam rangkaian panjang Zaman Es yang terjadi selama tahun-tahun terakhir Pleistosen, dari sekitar 110.000 sampai 12.000 tahun yang lalu. Sundalandia meliputi Semenanjung Malaya di daratan Asia, serta pulau-pulau besar Kalimantan, Jawa dan Sumatera, dan pulau-pulau sekitarnya. Batas timur Sundalandia adalah Garis Wallace, diidentifikasi oleh Alfred Russel Wallace sebagai batas timur kisaran daratan fauna mamalia Asia, dan juga sebagai batas zona ekosistem Indomalaya dan Australasia. Pulau-pulau di sebelah timur garis Wallace dikenal sebagai Wallacea, dan dianggap sebagai bagian dari Australasia. Perlu dicatat bahwa saat ini secara umum telah diterima bahwa Asia Tenggara adalah merupakan titik masuk migrasi manusia modern dari Afrika.
Nama "Sundalandia" pertama kali diusulkan oleh van Bemmelen pada tahun 1949, diikuti oleh Katili (1975), Hamilton (1979) dan Hutchison (1989), untuk menggambarkan sebuah inti benua Asia Tenggara yang membentuk bagian selatan lempeng Eurasia. Sundalandia berbatasan di sebelah barat, selatan dan timur dengan wilayah tektonik aktif yang ditandai dengan kegempaan dan aktivitas gunung berapi yang intensif. Zona tektonik aktif ini secara efektif merupakan sabuk pegunungan dalam proses pembentukannya, dan mengandung banyak fitur yang biasanya dianggap berhubungan dengan akresi pegunungan: terdapat subduksi aktif, transfer material pada batas lempeng, contoh tumbukan dengan fitur apung di lempeng samudera, busur dan benua dan banyaknya magmatisme.
Sabuk pegunungan yang ada sekarang terletak di persimpangan tiga lempeng utama: Lempeng Eurasia, Lempeng India, Lempeng Australia dan Lempeng Laut Pasifik-Filipina. Lempeng=lempeng ini mengelilingi Sundalandia dan membentang dari Sumatera ke Filipina melalui Indonesia timur. Karakteristik dan lebarnya berubah dari barat ke timur dan terdiri dari segmen-segmen yang berbeda atau jahitan-jahitan dengan karakter yang berbeda.

Gambar 1 – Peta Sundalandia
Laut Tiongkok Selatan dan daratan di sekitarnya telah diteliti oleh para ilmuwan seperti Molengraaff dan Umbgrove, yang mendalilkan sistem drainase kuno yang sekarang terendam. Daerah ini telah dipetakan oleh Tjia pada tahun 1980 dan dijelaskan secara lebih rinci oleh Emmel dan Curray pada tahun 1982 termasuk delta sungai, dataran banjir dan rawanya. Ekologi Paparan Sunda yang terekspos telah diselidiki dengan menganalisis inti bor dasar laut. Serbuk sari yang ditemukan didalam inti telah menunjukkan ekosistem yang kompleks dan berubah dari waktu ke waktu. Penggenangan Sundalandia telah memisahkan spesies yang pernah menempati lingkungan yang sama seperti ikan surau air tawar (Polydactylus macrophthalmus, Bleeker 1858; Polynemus borneensis, Vaillant 1893) yang pernah berkembang dalam sistem sungai yang sekarang disebut "Sungai Sunda Utara" atau "Sungai Molengraaff". Ikan ini sekarang ditemukan di Sungai Kapuas di pulau Kalimantan, serta di Sungai Musi dan Batanghari di pulau Sumatera.
Periode Glasial Terakhir, atau Zaman Es Terakhir, dianggap oleh para ilmuwan sebagai peristiwa glasiasi (pencairan es) terbaru dalam serangkaian Zaman Es yang lebih panjang, yang dimulai lebih dari dua juta tahun yang lalu dan telah mengalami beberapa glasiasi. Dalam periode ini, ada beberapa perubahan diantara gletser maju dan mundur. Glasiasi terluas dalam Periode Glasial Terakhir ini adalah sekitar 21.000 tahun yang lalu. Pola umum pendinginan dan gletser maju secara global adalah mirip, namun perbedaan lokal pengembangan gletser maju dan mundur dari benua ke benua yang terinci sulit untuk dibandingkan.
Dari sudut pandang arkeologi manusia, periode tersebut masuk kedalam periode Paleolitik dan Mesolitik. Saat peristiwa glasiasi dimulai, Homo sapiens hanya terdapat di Afrika dan menggunakan peralatan yang sebanding dengan yang digunakan oleh manusia Neanderthal di Eropa, Levant dan Homo erectus di Asia. Menjelang akhir peristiwa tersebut, Homo sapiens menyebar ke Eropa, Asia, dan Australia. Gletser mundur memungkinkan kelompok orang Asia bermigrasi ke Amerika dan mempopulasikannya.

Gambar 2 – Permukaan laut Pasca-Glasial
Stadial Dryas Muda, juga disebut Pendinginan Besar, adalah suatu kondisi geologi beriklim dingin dan kering yang singkat (1300 ± 70 tahun) dan terjadi antara sekitar 12.800 dan 11.600 tahun yang lalu. Stadial Dryas Muda diduga disebabkan oleh runtuhnya lapisan es di Amerika Utara, meskipun teori lain juga telah diusulkan. Stadial ini terjadi setelah interstadial Bølling-Allerød (periode hangat) pada akhir Pleistosen dan mendahului kelahiran Holosen Awal. Nama stadial ini diambil dari nama sebuah genus indikator, Dryas octopetala, bunga liar yang terdapat di tundra Pegunungan Alpin.
Stadial-stadial Dryas adalah periode-periode dingin yang menyelingi tren pemanasan sejak Maksimum Glasial Terakhir 21.000 tahun yang lalu. Dryas Tua terjadi sekitar 1.000 tahun sebelum Dryas Muda dan berlangsung sekitar 400 tahun. Dryas Tertua terjadi antara sekitar 18.000 dan 14.700 tahun yang lalu.

Gambar 3 – Suhu periode Glasial Terakhir berdasarkan pengukuran lapisan es di Greenland

Gambar 4 – Animasi suhu pada Maksimum Glasial Terakhir

Lihat di youtube
Batimetri dan Topografi

Gambar 5 – Sebuah peta yang menunjukkan Sundalandia pada sekitar Zaman Es Terakhir (21.000 tahun yang lalu) yang dihasilkan oleh penulis dari grid elevasi GTOPO30 yang diterbitkan oleh USGS. Muka air lautnya adalah sekitar 120 meter dibawah sekarang. Pola aliran sungai yang dibawah muka air laut sekarang digambar menggunakan grid tersebut dan perkiraan sedimentasi laut, transportasi sedimen pesisir, pembentukan delta, peristiwa meander, perubahan rezim sungai dan gerakan sungai. Sungai daratan sekaran digabungkan. Warna-warna selain biru menunjukkan elevasi permukaan tanah. Garis merah tipis adalah garis pantai sekarang.
Data topografi dan batimetri saat ini yang meliputi Paparan Sunda dalam proyeksi geografi (lintang dan bujur) telah diekstrak dari grid elevasi GTOPO30 yang diterbitkan oleh USGS. GTOPO30 meliputi resolusi spasial lintang dan bujur horizontal 30 detik busur (sekitar 0,9 km di dekat khatulistiwa) dalam bentuk berkas format model elevasi digital (digital elevation model, DEM). Grid serupa lainnya seperti GEBCO_8 yang diterbitkan oleh IHO dan IOC/UNESCO, dan ETOPO1 yang diterbitkan oleh NOAA juga digunakan sebagai referensi. Diterapka skema warna pada DEM dimana daerah dibawah -120 m diwakili oleh warna biru sehingga garis pantai pada periode Maksimum Glasial Terakhir dapat dengan mudah diidentifikasi.
Beberapa asumsi telah diterapkan dalam prosedur analitisnya (Sathiamurthy et al, 2006). Pertama, diasumsikan bahwa topografi dan batimetri daerah tersebut mendekati fisiografi yang ada selama rentang waktu dari 21.000 tahun yang lalu sampai sekarang. Namun, karena proses sedimentasi dan erosi telah mempengaruhi batimetri Paparan Sunda selama 21.000 tahun terakhir (Schimanski dan Stattegger, 2005), maka hasilnya adalah hanya perkiraan. Dengan demikian, harus ditekankan bahwa kedalaman dan geometri Paparan Sunda dan depresi-depresi yang sekarang terendam tidak mencerminkan kondisi masa lalu yang tepat.
Kedua, diasumsikan bahwa dasar laut yang sekarang adalah seperti yang ada pada periode Maksimum Glasial Terakhir dan sedikit dipengaruhi oleh erosi arus dasar laut, pelarutan kapur atau gerakan tektonik, seperti ditunjukkan oleh Umbgrove (1949), yang mungkin terjadi selama transgresi pada masa Pasca-Pleistosen awal. Dalam hal gerakan tektonik, Geyh et al (1979) menyebutkan bahwa Selat Sumatera adalah secara tektonik stabil setidaknya selama Holosen. Selanjutnya, Tjia et al (1983), menyatakan bahwa Paparan Sunda telah sebagian besar secara tektonik stabil sejak awal Tersier. Namun demikian, Tjia et al (1983) menunjukkan bahwa kenaikan permukaan laut di wilayah ini dapat dikaitkan dengan kombinasi dari kenaikan permukaan laut yang sebenarnya dan gerakan kerak vertikal. Hill (1968) mengacu pada karya sebelumnya yang dilakukan oleh Umbgrove (1949), menyarankan kemungkinan solusi kapur sebagai modus pembentukan depresi (seperti dalam kasus lubang Lumut lepas pantai Perak, Malaysia), dan memberikan alternatif penjelasan, yaitu disebabkan oleh proses tektonik.
Data sedimentasi dasar laut adalah jarang tersedia namun pendekatan proses sedimentasi telah dibuat dalam menghasilkan peta topografi dan batimetri daerah Sundalandia. Dalam kondisi yang sama, proses-proses lain seperti transportasi sedimen pesisir, pembentukan delta, proses meander, perubahan rezim sungai dan gerakan sungai juga telah didekati dan dimasukkan pada peta. Danau kuno direkonstruksi dari DEM dan riwayat geologi yang ada. Pulau-pulau yang kecil dan tidak signifikan telah dihapus.
Selain peta topografi dan batimetri, garis pantai pada permukaan laut tertentu, kemiringan permukaan tanah, daerah aliran sungai dan pola aliran sungai juga dihasilkan dan menempatkannya pada layer-layer yang berbeda.

Gambar 6 – Sebuah peta yang menunjukkan DAS utama Sundalandia di sekitar periode Maksimum Glasial Terakhir (21.000 tahun BP) yang dihasilkan oleh penulis menggunakan metode yang sama seperti pada gambar sebelumnya. Nama-nama sungai diberikan berdasarkan pada nama-nama laut, selat, teluk, pulau atau sungai saat ini yang ditempati oleh DAS.
Vegetasi
Cannon et al (2009) telah melakukan penelitian terhadap distribusi vegetasi di wilayah Sundalandia pada Maksimum Glasial Terakhir menggunakan model spasial eksplisit yang digabungkan dengan bukti-bukti geografi, paleoklimatologi dan geologi. Vegetasinya dibagi menjadi 3 macam, yaitu vegetasi pesisir dan rawa, vegetasi daerah rendah dan vegetasi daerah tinggi.
Vegetasi pesisir dan rawa mengalami sejarah biogeografi yang paling dinamis diantara ketiga macam vegetasi yang diteliti. Pada puncak Maksimum Glasial Terakhir, ketika permukaan laut turun dibawah garis pantai, hutan bakau terdapat didalam daerah yang sangat sempit di sepanjang pesisir. Namun, banyak rawa pesisir yang meluas sampai ke pedalaman ditumbuhi kerapah gambut dan kerangas yang berbagi dengan hutan rawa gambut pesisir. Ketika daratan mulai tergenangi, pada 11.000 – 9.000 tahun lalu, vegetasi pesisir dan rawa mengalami ekspansi yang dramatis tetapi relatif singkat. Pada sekitar 8.000 tahun lalu, vegetasi pantai telah kira-kira pada posisi yang sama dengan saat ini, dengan luasan bakau, aluvial air tawar dan rawa gambut ditentukan oleh pola progradasi masing-masing delta sungai yang mengikuti transgresi masa Holosen. Vegetasi pesisir dan rawa juga mengalami relokasi geografi yang tiba-tiba dan menyeluruh sampai ratusan kilometer selama penggenangan yang cepat.
Luasan vegetasi daerah rendah mencapai maksimum pada Maksimum Glasial Terakhir yang disebabkan oleh adanya daratan yang luas karena penurunan permukaan laut. Luasan vegetasi daerah tinggi juga mencapai maksimum pada Maksimum Glasial Terakhir. Secara umum, distribusi vegetasi daerah tinggi sangat sensitif terhadap interaksi antara perubahan suhu dan tingkat pertumbuhan vegetasi.

Gambar 7 – Peta vegetasi Sundalandia pada Maksimum Glasial Terakhir berdasarkan data historis dari Bird et al (2005) dan beberapa penyesuaian, dengan skenario koridor terbuka (kiri) dan tertutup (kanan) (Cannon et al, 2009)

Lihat di youtube.
Kondisi Saat ini
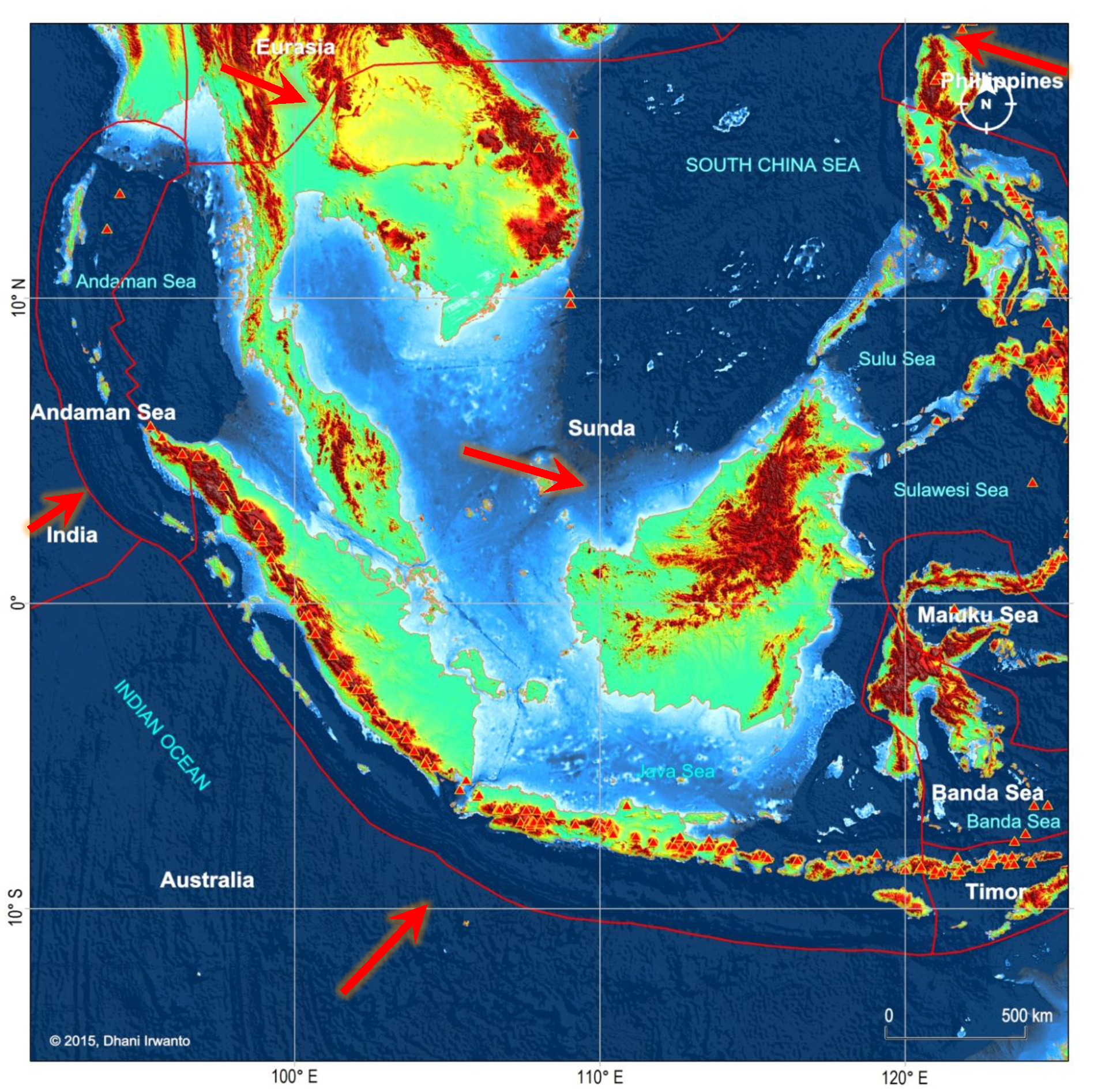
Gambar 8 – Sesar aktif utama di Sundalandia pada zona konvergensi Lempeng Sunda, Eurasia, Filipina, India dan Australia. Lempeng Timor dan Laut Banda yang lebih kecil (bagian dari Lempeng Sunda), Maluku (bagian dari Lempeng Filipina) dan Andaman (bagian dari Lempeng Eurasia) juga ditampilkan. Panah besar menunjukkan gerakan mutlak lempeng. Segitiga merah adalah gunungapi.

Gambar 9 – Plot kejadian gempa besar yang pernah tercatat beserta intensitasnya dalam skala Mw. Perhatikan bahwa Sundalandia dikelilingi oleh deretan rawan gempa. (Sumber: USGS)

Gambar 10 – Plot sumber tsunami yang pernah tercatat beserta akibat kenaikan airnya. Perhatikan bahwa tsunami sering terjadi di Laut Banda dan Laut Sulawesi yang dapat mempengaruhi pulau-pulau bagian dalam. (Sumber: NOAA)

Gambar 11 – Plot letusan gunungapi yang pernah diketahui beserta intensitasnya dalam Volcanic Explosivity Index (VEI). Perhatikan letusan skala besar Tambora pada tahun 1815 dan letusan-letusan Krakatau yang sering terjadi, terbesar pada tahun 1883. (Sumber: NOAA)
Migrasi Manusia
Menurut teori sebelumnya, nenek moyang masyarakat Austronesia modern di kepulauan Melayu dan wilayah yang berdekatan diyakini telah bermigrasi ke selatan, dari daratan Asia Timur ke Taiwan, dan kemudian ke seluruh kepulauan Asia Tenggara. Namun, temuan terakhir menunjukkan bahwa Sundalandia yang sekarang terendam adalah merupakan awalmula peradabah: disebut sebagai teori "Keluar dari Sundalandia".
Stephen Oppenheimer menempatkan asal Austronesia di Sundalandia dan daerah-daerah diatasnya. Penelitian genetik yang dilaporkan pada tahun 2008 menunjukkan bahwa pulau-pulau yang merupakan sisa-sisa Sundalandia kemungkinan besar dihuni sedini 50.000 tahun yang lalu, bertentangan dengan hipotesis sebelumnya (Bellwood dan Dizon, 2005) bahwa daerah tersebut dihuni paling lambat 10.000 tahun yang lalu dari Taiwan.
Sebuah studi oleh Universitas Leeds yang dipublikasikan dalam Molecular Biology and Evolution pada tahun 2008, yang meneliti garis keturunan DNA mitokondria, menunjukkan bahwa manusia telah mendiami pulau-pulau di Asia Tenggara dalam jangka waktu yang lebih lama dari yang diyakini sebelumnya. Penyebaran penduduk tampaknya telah terjadi pada saat yang sama dengan naiknya permukaan laut, yang telah mengakibatkan migrasi dari Kepulauan Filipina ke utara menuju Taiwan dalam 10.000 tahun terakhir. Migrasi penduduk yang paling mungkin adalah karena didorong oleh perubahan iklim – sebagai efek tenggelamnya sebuah benua kuno. Naiknya permukaan laut dalam tiga pulsa besar mungkin telah menyebabkan banjir dan perendaman di Sundalandia, menciptakan Laut Jawa dan Laut Tiongkok Selatan, dan ribuan pulau yang membentuk Indonesia dan Filipina kini. Berubahnya permukaan laut menyebabkan manusia tersebut untuk menjauh dari kediaman pantai dan budaya mereka, dan berpindah ke pedalaman di seluruh Asia Tenggara. Migrasi paksa ini menyebabkan manusia untuk beradaptasi dengan lingkungan hutan dan pegunungan baru, mengembangkan pertanian dan domestikasi, dan menjadi pendahulu manusia masa depan di wilayah ini.
Penelitian dan studi oleh HUGO Pan-Asian SNP Consortium pada tahun 2009, yang dilakukan dalam dan antara populasi yang berbeda di benua Asia, menunjukkan bahwa keturunan genetik sangat berhubungan dengan kelompok-kelompok etnis dan bahasa. Terdapat peningkatan yang jelas dalam keragaman genetik dari utara ke selatan. Penelitian ini juga menunjukkan bahwa ada satu aliran migrasi utama manusia ke Asia yang berasal dari Asia Tenggara, bukan beberapa aliran dalam dua arah selatan-utara seperti yang diusulkan sebelumnya. Hal ini menunjukkan bahwa Asia Tenggara merupakan sumber geografi utama populasi Asia Timur dan Asia Utara. Populasi Asia Timur terutama berasal dari Asia Tenggara dengan kontribusi kecil dari kelompok Asia Tengah-Selatan. Penduduk pribumi Taiwan berasal dari Austronesia. Hal ini berlawanan dengan teori bahwa Taiwan merupakan "tanah air" leluhur bagi populasi di seluruh Indo-Pasifik yang berbicara bahasa Austronesia.

Gambar 12 – Panah-panah berwarna yang menggambarkan meningkatnya diversifikasi genetik manusia setelah bermigrasi ke timur di sepanjang yang sekarang disebut pantai India dan berpencar menjadi beberapa kelompok genetik yang berbeda yang bergerak di seluruh Asia Tenggara dan bermigrasi ke utara ke Asia Timur (Sumber: HUGO Pan-Asian SNP Consortium)
Pada tahun 2012, Stephen Oppenheimer menunjukkan, berdasarkan bukti genetik, iklim dan arkeologi, bahwa manusia modern dari Afrika menyebar melalui jalur tunggal ke Sundalandia. Semua kelompok non-Afrika saat ini adalah keturunan dari penyebaran ini, dengan pengecualian beberapa autosom (7% atau kurang) yang berasal dari campuran dengan beberapa kelompok non-Afrika kuno. Terbukti bahwa terdapat penyebaran cepat melalui pantai Samudera Hindia ke Kalimantan dan Bali di ujung Paparan Sunda. Permukaan laut yang rendah pada Zaman Es memungkinkan penyebaran berikutnya ke Paparan Sahul, yang kemudian terisolasi cukup lama karena naiknya permukaan laut sampai masa pasca-glasial dimana para pelayar dari kepulauan Asia Tenggara menemukannya.

Gambar 13 – Peta yang menunjukkan jalur migrasi selatan tunggal dari Afrika dan jalur migrasi pantai dari Laut Merah di sepanjang pantai Indo-Pasifik ke Australia, termasuk kecenderungan ekstensi ke Tiongkok, Jepang dan Papua. Vegetasi yang tampak adalah pada Maksimum Glasial Terakhir. (Sumber: Oppenheimer, 2013)
Lihat Jurney of Mankind oleh The Bradshaw Foundation.
Pada tahun 2012, Jinam et al menentukan 86 urutan genom DNA mitokondria (mtDNA) lengkap dari empat masyarakat adat Malaysia, bersama-sama dengan analisis ulang terhadap data autosomal tunggal nukleotid polimorfisme (SNP) di Asia Tenggara untuk menguji kewajaran dan dampak model migrasi. Tiga kelompok Austronesia (Bidayuh, Selatar, dan Temuan) menunjukkan frekuensi tinggi haplogrup mtDNA, yang berasal dari daratan Asia 30,000–10,000 tahun lalu, tetapi terdapat frekuensi yang rendah penanda “Keluar dari Taiwan”. Analisis komponen utama dan analisis filogenetik menggunakan data SNP autosomal menunjukkan dikotomi antara kelompok Austronesia benua dan pulau. Mereka berpendapat bahwa baik mtDNA dan data autosomal menyarankan migrasi “Rangkaian Awal” yang berasal dari Indocina atau Tiongkok Selatan sekitar masa akhir-Pleistosen sampai awal-Holosen, yang mendahului, tetapi belum tentu mengecualikan, ekspansi Austronesia.
Karafet et al (2014), melalui sebuah studi Y-DNA, mendukung hipotesis bahwa Asia Tenggara adalah pusat diversifikasi garis keturunan haplogrup K di Oseania dan menggarisbawahi pentingnya potensi Asia Tenggara sebagai sumber variasi genetis untuk populasi Eurasia. Struktur filogenetis haplogrup K-M526 menunjukkan peristiwa percabangan yang berturut-turut (M526, P331 dan P295), yang tampaknya telah secara cepat terdiversifikasi. Kecuali P-P27, semua garis keturunannya saat ini terdapat Asia Tenggara dan Oseania. Struktur filogenetis haplogrup K-M526 dibagi menjadi empat subklade utama (K2a-d). Yang terbesar dari subklade ini, K2b, dibagi lagi menjadi dua kelompok: K2b1 dan K2b2. K2b1 menggabungkan haplogrup sebelumnya yang dikenal dengan M, S, K-P60 dan K-P79, sedangkan K2b2 terdiri dari haplogrup P dan sub haplogrupnya, Q dan R.
Menariknya, kelompok monofiletis yang dibentuk oleh haplogrup Q dan R, yang membentuk mayoritas garis keturunan patrilinial di Eropa, Asia Tengah dan Amerika, merupakan satu-satunya subklade K2b yang ternyata tidak terbatas secara geografis di Asia Tenggara dan Oseania saja. Perkiraan interval waktu peristiwa percabangan antara M9 dan P295 menunjukkan proses diversifikasi awal K-M526 yang cepat, yang kemungkinan terjadi di Asia Tenggara, yang kemudian mengekspansi ke arah barat menjadi nenek moyang haplogrup Q dan R. Menariknya lagi, bukti DNA purba menunjukkan bahwa haplogrup R1b – garis keturunan yang dominan saat ini di Eropa Barat – mencapai frekuensi yang tinggi setelah periode Neolitikum Eropa seperti yang ditunjukkan oleh Lacan et al dan Pinhasi et al.

Gambar 14 – Filogeni haplogrup K (Karafet et al, 2014)

Gambar 15 – Penyebaran keturunan haplogrup K2 (Karafet et al, 2014)
Brandão et al dalam sebuah makalah yang diterbitkan oleh Human Genetics pada tahun 2016 menyatakan bahwa penyebaran keramik Neolitik dan bahasa Melayu-Polinesia di kepulauan Asia Tenggara yang menunjukkan penyebaran pertanian keluar dari Taiwan 4.000 tahun yang lalu adalah masih menjadi perdebatan. Mereka kemudian melakukan studi menggunakan analisis DNA mitokondria (mtDNA) untuk mengidentifikasi kelompok garis keturunan utama yang paling mungkin telah tersebar dari Taiwan ke kepulauan Asia Tenggara, dan menyimpulkan bahwa penyebaran tersebut memiliki dampak yang relatif kecil pada struktur genetik yang masih ada di kepulauan Asia Tenggara, dan bahwa peran pertanian dalam perluasan bahasa Austronesia adalah kecil. Apabila dibandingkan antara penyebaran “Keluar dari Taiwan” pada pertengahan Holosen dengan penyebaran pasca-glasial dari kepulauan Asia Tenggara pada awal Holosen, hanya sekitar 20% garis keturunan mtDNA di kepulauan Asia Tenggara saja yang merupakan hasil penyebaran “Keluar dari Taiwan” dan selebihnya adalah hasil penyebaran pasca-glasial dari kepulauan Asia Tenggara, terutama karena naiknya permukaan laut pada Maksimum Glasial Terakhir. Migrasi dari Asia daratan ke Taiwan terjadi sekitar 6.000 – 7.000 tahun yang lalu dan tetap menjadi penduduk asli yang berbeda sampai dengan penyebarannya ke kepulauan Asia Tenggara.
Analisis rinci yang menggabungkan data DNA mitokondria (mtDNA), kromosom-Y dan genome dalam studi yang dilakukan oleh Soares et al dari University of Minho di Portugal dan diterbitkan dalam jurnal Human Genetics pada 2016, menunjukkan rangkaian peristiwa yang jauh lebih rumit. mtDNA dan kromosom-Y yang ditemukan di Kepulauan Pasifik telah ada di kepulauan Asia Tenggara jauh lebih awal dari 4.000 SM, yang menimbulkan keraguan yang serius terhadap teori “Keluar dari Taiwan”. Soares et al berpendapat bahwa lanskap dan permukaan laut yang berubah sekitar 11.500 tahun lalu menyebabkan ekspansi yang signifikan dari Indonesia 8.000 tahun lalu. Ekspansi ini, yang merupakan penemuan tim, menunjukkan bahwa populasi di seluruh Asia Tenggara dan Kepulauan Pasifik berbagi mtDNA dan kromosom-Y yang sama. Hasil studi tim tersebut juga menunjukkan adanya gelombang minor migrasi yang mungkin mengarah pada penyebaran bahasa Austronesia.

Gambar 16 – Garis keturunan ibu yang terlibat dalam migrasi manusia utama di wilayah Asia Tenggara dan Taiwan (Soares et al, 2016)


Edwina Palmer (2007) telah melakukan studi bahwa penggenangan Sundalandia pasca-glacial telah mendorong sebagian penduduknya untuk bermigrasi pada sekitar 10.000 – 11.000 tahun yang lalu, mengikuti sabuk hutan lucidophyllous yang meluas dan akhirnya menetap di Jepang sekarang pada periode Jomon. Orang-orang ini mungkin penutur bahasa Austronesia. Selanjutnya, ia berpendapat bahwa migrasi penutur bahasa Austronesia “Keluar dari Taiwan” bisa terjadi kemudian sebagai migrasi balik pada periode Holosen akhir, dan bahwa skenario migrasi “Keluar dari Sundalandia” ke Jepang pada periode Jomon belum tentu sepenuhnya kompatibel dengan teori “Keluar dari Taiwan” tersebut.
Migrasi dari Asia Tenggara ke Madagaskar telah dibuktikan dengan studi hubungan linguistik antara bahasa Madagaskar dengan bahasa Barito Kalimantan dan telah dikonfirmasi dengan penelitian genetik. Analisis genetik serupa juga telah diterapkan pada tanaman, dan dengan mempelajari terminologi nama-nama yang digunakan juga telah menunjukkan migrasi tersebut (Claude Allibert, 2011). Migrasi orang-orang Asia Tenggara ke Madagaskar berasal dari bagian selatan Sulawesi (Bugis) dan Kalimantan (Maanyan). Kemiripan karakteristik antropologi budaya dan agama, seperti keyakinan tentang kembaran anak dan hewan, dan juga praktek penguburan ganda (terdapat di Filipina), menguatkan indikator kaitan genetik dan linguistik mereka.

Gambar 17 – Peta yang menunjukkan lokasi peninggalan pra-sejarah di Sundalandia, yang dapat berupa batu megalitik (menhir), manusia prasejarah, seni cadas dan punden berundak. Perhatikan bahwa peninggalan tersebut banyak terdapat di Jawa, Bali, Sumatera Selatan, Sulawesi Selatan dan Kalimantan Selatan. Lokasi bawah laut belum ada yang diteliti. (Sumber: berbagai sumber, yang dikumpulkan oleh penulis)
Peradaban Sungai
Sungai menyediakan aliran dan pasokan air yang terus menerus atau yang selalu dapat diandalkan untuk transportasi, pertanian dan bahan makanan bagi manusia. Sungai-sungai tersebut bersama dengan iklim, vegetasi, geografi dan topografi menjadikan awal berkembangannya peradaban sungai. Selain itu, sementara masyarakat dari peradaban tersebut bergantung pada sungai, sungai juga menjadi tempat awalmulanya inovasi baru dan perkembangan teknologi, ekonomi, kelembagaan dan organisasi. Budaya sungai adalah tempat lahirnya peradaban maritim yang kemudian berkembang menjadi masyarakat penutur bahasa Austronesia.
Sungai-sungai besar dengan lahan yang subur terdapat di Sundalandia pada Zaman Es. Besar kemungkinan bahwa peradaban di Sundalandia berawal di sungai-sungai tersebut. Karena laut juga tidak dapat terpisahkan dari kehidupan mereka, maka tumbuhnya peradaban hingga menjadi besar adalah di sekitar muara sungai. Kenaikan permukaan laut dan bencana banjir atau tsunami yang berulang-ulang menyebabkan mereka untuk berpindah ke tempat yang lebih tinggi, di pegunungan. Sarana transportasi yang ada pada waktu itu hanyalah sungai, sehingga mereka berpindah ke daerah-daerah hulu sungai. Peradaban kuno dan yang masih berlanjut sampai sekarang telah diamati dan mereka berada di daerah-daerah hulu sungai besar.

Gambar 18 – Peradaban sungai di Sundalandia
Pembudidayaan
Penelitian-penelitian yang sudah ada menunjukkan bahwa pembudidayaan beberapa tanaman pertanian dan hewan peliharaan pertama kali dilakukan di Sundalandia dan wilayah-wilayah sekitarnya yang erat kaitannya dengan penyebaran penduduk dari Sundalandia. Namun demikian, penelitian-penelitian tersebut terbatas pada penemuan-penemuan yang ada pada saat ini saja. Di Sundalandia, kedua pembudidayaan tersebut tidak terlepas dari lingkungan air, baik itu sungai maupun laut, yang keduanya bertemu di muara sungai. Dengan demikian dapat diduga bahwa pusat-pusat peradaban awal adalah dimulai dari lingkungan muara sungai, seperti telah dibahas diatas. Namun, muara-muara sungai besar di Sundalandia pada masa Zaman Es Akhir saat ini berada dibawah permukaan laut. Dapat diduga bahwa bukti-bukti pembudidayaan yang paling tua belum dapat ditemukan karena berada di dasar laut dan bukti-bukti yang ada saat ini adalah di daratan yang lebih tinggi sehingga usianya lebih muda. Selain itu, Sundalandia memiliki aktifitas gunung berapi yang tinggi sehingga daratan yang ada saat ini telah tertutup abu volkanik yang sangat tebal, yang menjadi hambatan yang cukup serius untuk menemukan bukti-bukti arkeologinya.
Kelapa
Analisis DNA terhadap lebih dari 1.300 buah kelapa dari seluruh dunia mengungkapkan bahwa kelapa (Cocos nucifera) pada awalnya dibudidayakan di dua lokasi terpisah, yaitu di sekitar Samudera Pasifik dan Hindia (Baudouin et al, 2008; Olsen et al, 2011). Selain itu, genetika kelapa juga tercatat dalam rute perdagangan pra-sejarah dan kolonisasi Amerika. Di sekitar Samudera Pasifik, kelapa pertama kali dibudidayakan di kepulauan Asia Tenggara, yaitu Filipina, Malaysia, Indonesia, dan mungkin juga di daratan Asia. Di sekitar Samudera Hindia, kemungkinan pusat budidayanya adalah pinggiran selatan India, termasuk Srilanka, Maladewa dan Lakadewa. Kelapa dari sekitar Samudera Pasifik diperkenalkan ke sekitar Samudera Hindia beberapa ribu tahun lalu oleh masyarakat penutur Austronesia kuno yang membangun jalur perdagangan yang menghubungkan Asia Tenggara dengan Madagaskar dan pesisir Afrika timur.
Baca juga: Kelapa
Padi
Dalam buku Eden in the East (1998), Stephen Oppenheimer mengklaim bahwa budidaya padi bukan dimulai di Tiongkok tetapi di Semenanjung Malaya, 9.000 tahun lalu. Butiran beras telah ditemukan, yang berusiakan antara 7.000 sampai 5.000 SM, di Semenanjung Malaya. Periode ini adalah beberapa ribu tahun lebih tua dari kedatangan orang-orang Austronesia dari Taiwan yang diduga telah membawa teknologi pertanian ke Asia Tenggara.
Ada empat varietas utama padi: japonica, berbutir pendek yang tumbuh di Jepang, Korea dan Tiongkok timur; indica, berbutir panjang yang umum di India, Pakistan dan sebagian besar Asia Tenggara; aus, tumbuh terutama di Banglades; dan beras wangi, yang meliputi varietas yang lebih eksotis seperti basmati di India dan melati di Thailand. Para ilmuwan terutama memfokuskan pada indica dan japonica karena temuan arkeologi menunjukkan keduanya memiliki sejarah budidaya yang panjang. Para peneliti umumnya sepakat bahwa japonica telah dibudidayakan di Tiongkok selatan atau Asia Tenggara antara 8.200 dan 13.500 tahun lalu. Lokasi yang tepat masih diperdebatkan.
Para ahli masih memperdebatkan asal pembudidayaan indica. Peter Civáň dari University of Manchester, Inggris dan timnya pada 2015, dengan membandingkan DNA dari 1.083 varietas padi modern dengan 446 sampel padi yang diambil dari seluruh Asia Selatan, telah melacak kembali sejarah tanaman tersebut menjadi tiga jenis padi yang berbeda. Japonica, yang disukai di Jepang, ditelusuri kembali dari tengah Lembah Yangtze di Tiongkok Selatan. Tim tersebut telah melacak kembali kelompok indica ke lembah Sungai Brahmaputra yang mengalir dari Himalaya, sementara kelompok aus berasal dari wilayah yang sekarang India dan Banglades.
Namun demikian, penelitian tentang asal pembudidayaan padi masih terus berjalan. Dapat diduga bahwa bukti-bukti budidaya padi yang paling tua belum dapat ditemukan karena berada di dasar laut dan bukti-bukti yang ada saat ini adalah di daratan yang lebih tinggi sehingga usianya lebih muda. Bukti-bukti di daratan juga belum tentu mencerminkan kondisi yang sebenarnya karena daerah Sundalandia adalah pada umumnya tertutup oleh abu volkanik yang sangat tebal.n.
Pisang
Pisang (Musa spp) diyakini berasal lebih dari 10.000 tahun yang lalu dan beberapa ilmuwan percaya bahwa pisang adalah buah-buahan yang pertama kali dibudidayakan. Pisang yang ada saat ini jauh lebih baik daripada buah liar asli yang berisi banyak biji yang besar dan keras dan rasanya kurang enak. Dua varietas pisang liar, Musa acuminata dan Musa baalbisiana telah dipersilangkan sehingga menghasilkan pisang tanpa biji seperti yang ada saat ini.
Pisang diperkirakan pertama kali tumbuh di daerah yang mencakup Semenanjung Malaya, Indonesia, Filipina dan Papua. Dari sini, para pedagang dan penjelajah membawanya ke India, Afrika dan Polinesia. Sebuah kitab Buddha, dikenal sebagai Pali Canon, kira-kira pada 600 SM, mencatat pedagang India yang melakukan perjalanan melalui wilayah Melayu telah memakan buah tersebut dan membawa pulang tanaman itu. Pada 327 SM, ketika Alexander Agung dan pasukannya menyerbu India, ia mendapati tanaman pisang di lembah India. Setelah mencicipi buah yang tidak biasa ini untuk pertama kalinya, ia memperkenalkan penemuan baru ini kepada dunia Barat.
Pisang telah menyebar ke Tiongkok pada sekitar 200 M. Menurut sejarawan Tiongkok Yang Fu, pisang hanya pernah tumbuh di wilayah Tiongkok selatan. Pisang di Tiongkok tidak pernah benar-benar populer sampai abad ke-20 karena dianggap sebagai buah yang asing, aneh dan eksotis. Pisang mulai dikembangkan di Afrika sekitar 650 M.
Diperkirakan bahwa pedagang dari Arab, Persia, India dan Indonesia mendistribusikan pisang ke sekitar daerah pesisir Samudera Hindia antara abad ke-5 dan ke-15. Pelaut Portugis mendapatkan pisang di Afrika Barat dan perkebunan pisang didirikan pada abad ke-15 di lepas pantai negaranya, di Kepulauan Kanaria. Antara abad ke-16 dan ke-19, pisang diperdagangkan di Amerika dan perkebunan-perkebunan didirikan di Amerika Latin dan Karibia. Tanaman pisang pertama kali tiba di Australia pada tahun 1800-an.
Tebu
Tebu (Saccharum spp) kemungkinan pertama kali dibudidayakan oleh masyarakat Papua, sekitar 8.000 SM. Namun, teknik ekstraksi dan teknologi pemurniannya lalu dikembangkan oleh orang-orang yang tinggal di India. Setelah pembudidayaan tersebut, kemudian menyebar dengan cepat ke Asia Tenggara dan Tiongkok Selatan. Di India, dimana proses penyulingan sari tebu menjadi kristal butiran dikembangkan, sering dikunjungi oleh konvoi kekaisaran (seperti dari Tiongkok) untuk belajar tentang budidaya dan penyulingan tebu. Pada abad ke-5, budidaya tebu dan pengolahannya telah mencapai Persia; dan dari sana pengetahuan tersebut dibawa ke Mediterania oleh ekspansi Arab.
Eksplorasi dan penaklukan oleh Spanyol dan Portugis pada abad ke-15 membawa tebu ke baratdaya Iberia. Henry the Navigator memperkenalkan tebu ke Madeira pada 1425, sementara Spanyol, yang akhirnya dapat menaklukkan Kepulauan Kanaria, memperkenalkan tebu disana. Pada 1493, dalam perjalanan kedua, Christophorus Columbus membawa bibit tebu ke Amerika, khususnya Hispaniola.
Cabai
Penelitian yang ada menunjukkan bahwa cabai (Capsicum spp) dibudidayakan lebih dari 6.000 tahun lalu di Meksiko, di wilayah yang meluas dari Puebla Selatan dan Oaxaca Utara sampai ke Veracruz Tenggara, dan merupakan salah satu tanaman dengan penyerbukan sendiri yang dibudidayakan di Meksiko. Namun, cabai telah disebutkan dalam Siva Purana dan Wamana Purana, di India, yang berusiakan sekitar abad ke-6 sampai ke-8 M (Banerji, 1980). Nama Sansekerta-nya adalah marichi-phalam yang diterapkan untuk Capsicum annuum dan Capsicum frutescens (Nadkarni, 1914). Tanaman dan buahnya secara natural digambarkan dalam ukiran batu di sebuah candi Siwa di Tiruchirapalli, Tamil Nadu (Gupta, 1996). Sebuah ukiran tanaman cabai yang sangat eksplisit ditemukan pada panel dinding reruntuhan di kompleks Candi Prambanan, Jawa Tengah, yang berusiakan seribu tahun lebih.
Jagung
Penelitian yang ada menunjukkan bahwa jagung (Zea mays) dibudidayakan kurang lebih 10.000 tahun lalu oleh orang asli Meksiko. Namun, penyelidikan lapangan telah menemukan jenis jagung yang tidak biasa yang tumbuh di Asia (terutama di suku Sikkim di pedalaman Himalaya dan varietas “berlilin” di Myanmar, seluruh Tiongkok dan Semenanjung Korea), sebagian besar jauh dari daerah pesisir. Karakteristik dan distribusi jagung ini tidak dapat dijelaskan pada masa pasca-Columbus, karena varietas berlilin tidak dikenal di Amerika. Johannessen et al (1998a, 1989a) telah mendokumentasikan secara luas bahwa ukiran jagung – ratusan jumlahnya – terdapat pada dinding candi di Karnataka, India selatan. Seni ini biasanya berasal dari abad ke-11 sampai ke-13 M, tetapi beberapa buah adalah jauh lebih tua. Empat kata Sansekerta untuk jagung telah teridentifikasi, sedangkan Garuda Purana dan Linga Purana di India (abad ke-5 M) telah mencatat tentang jagung. Sebuah keramik yang ditemukan di Zhenghou, Tiongkok yang berusiakan sekitar 2.000 tahun menunjukkan bekas jagung yang tercetak sebelum dibakar. Sebuah ukiran tanaman jagung ditemukan pada panel dinding reruntuhan di kompleks Candi Prambanan, Jawa Tengah, di samping ukiran cabai, yang berusiakan seribu tahun lebih.
Ayam
Hasil analisis DNA purba yang dilakukan oleh Alice A Storey et al pada 2012 terhadap 48 tulang ayam yang berasal dari data arkeologi memberikan petunjuk tentang penyebaran ayam ternak oleh manusia pra-sejarah. Tanda genetis haplogrup E mtDNA menghasilkan petunjuk bahwa ayam terdapat di Eropa pada 1,000 tahun lalu dan di sekitar Pasifik pada 3.000 tahun lalu, yang menunjukkan beberapa dispersal pra-sejarah dari Asia. Kedua jalur penyebaran berkumpul di Amerika dimana ayam diperkenalkan oleh masyarakat Polinesia dan kemudian oleh orang Eropa.
Penelitian yang dilakukan oleh Martin Johnsson di Department of Physics, Chemistry and Biology Linköping University, Swedia pada 2015 menunjukkan bahwa ayam pertama kali dibudidayakan dari bentuk liarnya yang disebut ayam hutan merah (Gallus gallus), suatu spesies ayam yang masih terdapat secara liar di sebagian besar Asia Tenggara, kemungkinan dihibridisasi dengan ayam hutan abu-abu (Gallus sonneratii), yang dilakukan mungkin sekitar 8.000 tahun lalu. Penelitian tersebut menunjukkan kemungkinan adanya beberapa asal daerah yang berbeda di Asia Tenggara dan Selatan, termasuk Tiongkok Selatan dan Utara, Thailand, Myanmar dan India.
Anjing
Penelitian yang dilakukan oleh Mattias Oskarsson di School of Biotechnology, Royal Institute of Technology (KTH), Swedia pada 2012 menggunakan urutan DNA kromosom-Y menunjukkan bahwa anjing di Asia Tenggara di sebelah selatan Sungai Yangtze memiliki keragaman genetis yang tertinggi dan diturunkan dari sejumlah besar keturunan serigala. Ia menekankan bahwa penyebaran anjing pada masa awal adalah erat hubungannya dengan sejarah manusia dengan anjing sebagai bagian budaya mereka. Ia untuk pertama kalinya menyelidiki penyebaran anjing dari Asia Tenggara ke Polinesia dan Australia, dan hasilnya dapat digunakan sebagai bukti untuk menelusuri asal-usul masyarakat Polinesia yang telah sebelumnya terindikasikan dari studi arkeologi dan linguistis.
Peter Savolainen dari KTH-Royal Institute of Technology di Swedia dan Ya-Ping Zhang dari Kunming Institute of Zoology di Tiongkok pada 2015 secara bersamaan menunjukkan bahwa manusia pertama kali menjinakkan anjing di Asia Tenggara 33.000 tahun lalu, dan bahwa sekitar 15.000 tahun lalu subset nenek moyang anjing mulai bermigrasi kearah Timur Tengah dan Afrika. Penyebaran tersebut mungkin terinspirasi oleh sifatnya yang dapat bersahabat dengan manusia, tetapi mungkin juga bahwa mereka melakukan perjalanannya secara mandiri. Salah satu faktor pendorong yang mungkin adalah mencairnya gletser, yang mulai mundur sekitar 19.000 tahun lalu. Tidak sampai 5,000 tahun setelah mereka pertama kali mulai menyebar dari Asia Tenggara, anjing diduga telah mencapai Eropa. Sebelum akhirnya mencapai Amerika, salah satu kelompok di Asia telah berasimilasi dengan anjing yang telah bermigrasi kembali ke Tiongkok Utara.
Babi
Bukti arkeologi yang ada saat ini menunjukkan bahwa babi awalnya dibudidayakan setidaknya di dua tempat, di lembah Mekong dan di Anatolia, wilayah di Turki kini. Studi yang dilakukan pada 2007 terhadap materi genetis dari 323 babi modern dan 221 babi kuno di Eurasia Barat menunjukkan bahwa babi pertama kali datang ke Eropa dari Timur Dekat, tetapi Eropa kemudian menjinakkan babi hutan lokal, yang tampaknya menggantikan babi asli mereka.
Penelitian yang dilakukan oleh Laurent Frantz dari University of Oxford, Inggris dan Martien Groenen dari Wageningen University and Research Centre, Belanda beserta timnya menggunakan analisis komputer canggih terhadap 103 urutan genom keturunan babi hutan dan babi peliharaan dari seluruh Eropa dan Asia, yang diterbitkan dalam Nature Genetics pada 2015, menunjukkan bahwa babi memang berasal dua tempat berbeda tersebut. Tetapi babi Eropa moderen adalah campuran yang berasal dari beberapa populasi babi hutan. Beberapa materi genetis mereka tidak dapat ditemukan pada DNA babi hutan yang dikumpulkan oleh para peneliti tersebut, sehingga mereka berkesimpulan bahwa setidaknya nenek moyang mereka berasal dari salah satu kelompok yang telah punah atau dari kelompok lain di Asia Tenggara/Timur. Anomali ini menunjukkan bahwa babi telah dibawa dari satu tempat ke tempat lainnya, dimana mereka diasimilasikan dengan kelompok tersebut.
Gajah Kalimantan
Asal gajah Kalimantan (Elephas maximus borneensis) adalah kontroversial. Dua hipotesis yang bersaing berpendapat bahwa mereka adalah asli dari masa Pleistosen, atau dibawa dari tempat lain. Secara taksonomi, mereka telah diklasifikasikan sebagai subspesies yang unik atau ditempatkan dibawah subspesies India atau Sumatera. Prithiviraj Fernando et al pada 2003 telah melakukan penelitian terhadap DNA mitokondria gajah Kalimantan dan gajah yang terdapat di seluruh Asia. Ia menemukan bahwa gajah Kalimantan secara genetis adalah berbeda, dengan indikasi molekul yang divergen pada kolonisasi masa Pleistosen sekitar 300.000 tahun lalu. Pada waktu kenaikan permukaan laut di Zaman Es terakhir yang memisahkan Pulau Kalimantan dari daratan Asia, gajah di pulau ini menjadi terisolasi dari sepupu mereka di daratan Asia dan Sumatera dan kemudian berkembang menjadi sub-spesies gajah Asia yang berbeda. Gajah Jawa (Elephas maximus sondaicus) yang sekarang telah punah adalah identik dengan gajah Kalimantan.
Teori Atlantis di Sundalandia
Beberapa penulis telah secara khusus menyatakan hubungan yang jelas antara Sundalandia dan Atlantis-nya Plato. Dataran bawah laut Sunda adalah cukup cocok dengan deskripsi Plato tentang Atlantis. Topografi, iklim, flora dan faunanya bersama-sama dengan aspek mitologi lokal, semuanya menjadi hal yang meyakinkan untuk mendukung ide ini.
CW Leadbeater (1854-1934), seorang teosopis ternama, adalah mungkin yang pertama menunjukkan adanya hubungan antara Atlantis dan Indonesia dalam bukunya, The Occult History of Java. Peneliti-peneliti lain telah menulis tentang prasejarah daerah tersebut, diantaranya yang paling dikenal adalah mungkin Stephen Oppenheimer yang dengan tegas menempatkan Taman Eden di wilayah ini, meskipun ia hanya menyebut sedikit referensi mengenai Atlantis. Robert Schoch, bekerjasama dengan Robert Aquinas McNally, menulis sebuah buku dimana mereka menunjukkan bahwa bangunan piramida mungkin memiliki asal-usul dari sebuah peradaban yang berkembang di bagian-bagian Sundalandia yang kini terendam.
Buku pertama yang secara khusus mengidentifikasi Sundalandia dengan Atlantis ditulis oleh Zia Abbas. Namun, sebelumnya telah ada setidaknya dua publikasi internet yang membahas secara rinci perihal Atlantis di Asia Tenggara. William Lauritzen dan almarhum Profesor Arysio Nunes dos Santos keduanya mengembangkan situs internet secara luas. Lauritzen juga telah menulis sebuah e-book yang tersedia dalam situsnya, sementara Santos mengembangkan pandangannya tentang Atlantis di Asia melalui bukunya, Atlantis: The lost continent finally found. Dr Sunil Prasannan membuat sebuah esai yang menarik didalam website Graham Hancock. Sebuah situs yang lebih esoteris juga menyampaikan dukungan mengenai teori Sundalandia.
Dukungan lebih lanjut tentang Atlantis di Indonesia terjadi pada April 2015 dengan penerbitan buku, Atlantis: The lost city is in Java Sea oleh seorang pakar hidrologi, Dhani Irwanto, yang berupaya untuk mengidentifikasi fitur kota yang hilang dalam rincian narasi Plato dengan suatu lokasi di Laut Jawa lepas pantai pulau Kalimantan.
Referensi
Stephen Oppenheimer, Out-of-Africa, the peopling of continents and islands: tracing uniparental gene trees across the map, Philosophical Transactions of The Royal Society B (2012) 367, 770–784
Andreia Brandão, Ken Khong Eng, Teresa Rito, Bruno Cavadas, David Bulbeck, Francesca Gandini, Maria Pala, Maru Mormina, Bob Hudson, Quantifying the legacy of the Chinese Neolithic on the maternal genetic heritage of Taiwan and Island Southeast Asia, Human Genetics, April 2016, Volume 135, Issue 4, pp 363-376
Tatiana M Karafet, Fernando L Mendez, Herawati Sudoyo, J Stephen Lansing and Michael F Hammer, Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia, European Journal of Human Genetics (2015) 23, 369–373
Pedro A Soares et al, Resolving the ancestry of Austronesian-speaking populations, Human Genetics Volume 135, Issue 3, pp 309-326, March 2016
Timothy A. Jinam, Lih-Chun Hong, Maude E Phipps, Mark Stoneking, Mahmood Ameen, Juli Edo, HUGO Pan-Asian SNP Consortium and Naruya Saitou, Evolutionary History of Continental Southeast Asians: “Early Train” Hypothesis Based on Genetic Analysis of Mitochondrial and Autosomal DNA Data, Society for Molecular Biology and Evolution 29(11):3513–3527, June 2012
Martin Johnsson, Genomics of chicken domestication and feralisation, IFM Biology, Department of Physics, Chemistry and Biology, Linköping University, Sweden, 2015
Storey AA, Athens JS, Bryant D, Carson M, Emery K, et al, Investigating the Global Dispersal of Chickens in Prehistory Using Ancient Mitochondrial DNA Signatures, PLoS ONE 7(7): e39171, 2012. doi:10.1371/journal.pone.0039171
Mattias Oskarsson, Analysis of the origin and spread of the domestic dog using Y-chromosome DNA and mtDNA sequence data, Division of Gene Technology, School of Biotechnology, Royal Institute of Technology (KTH), Stockholm, Sweden, 2012
Peter Savolainen et al, Out of southern East Asia: the natural history of domestic dogs across the world, Cell Research 26:21-33, 2015 doi:10.1038/cr.2015.147
Laurent A F Frantz, Joshua G Schraiber, Ole Madsen, Hendrik-Jan Megens, Alex Cagan, Mirte Bosse, Yogesh Paudel, Richard P M A Crooijmans, Greger Larson & Martien A M Groenen, Evidence of long-term gene flow and selection during domestication from analyses of Eurasian wild and domestic pig genomes, Nature Genetics Volume 47 Number 10, Oktober 2015
Prithiviraj Fernando, TNC Vidya, John Payne, Michael Stuewe, Geoffrey Davison, Raymond J Alfred, Patrick Andau, Edwin Bosi, Annelisa Kilbourn, Don J Melnick, DNA Analysis Indicates That Asian Elephants Are Native to Borneo and Are Therefore a High Priority for Conservation, PLoS Biology, Volume 1, Issue 1, 2003, pp 110 – 115
Peter Civáň, Hayley Craig, Cymon J Cox dan Terence A Brown, Three geographically separate domestications of Asian rice, Nature Plants 1, Article number: 15164, 2015, doi: 10.1038/ nplants.2015.164
Dhani Irwanto, Atlantis: Kota yang hilang ada di Laut Jawa, Indonesia Hydro Media, 2016
Hak cipta © 2015-2017, Dhani Irwanto
Berdasarkan naskah asli Sundaland

Buku ini menyajikan banyak indikasi dari data arkeologi, studi genetika, legenda, mitos, dan dongeng bahwa Sundalandia, sebuah wilayah bio-geografi Asia Tenggara yang terpapar selama periode Glasial Terakhir, adalah tempat lahir peradaban manusia.
Tidak ada komentar:
Posting Komentar